北京工商大学备课纸备注第九章细胞骨架第一节微丝微丝(microfilament,MF)是由肌动蛋白(actin)组成的直径约7nm的骨架纤维,又称肌动蛋白纤维actinfilament
微丝和它的结合蛋白(associationprotion)以及肌球蛋白(myosin)三者构成化学机械系统,利用化学能产生机械运动
一、分子结构根据等电点的不同可将高等动物细胞内的肌动蛋白分为3类,α分布于各种肌肉细胞中,β和γ分布于肌细胞和非肌细胞中
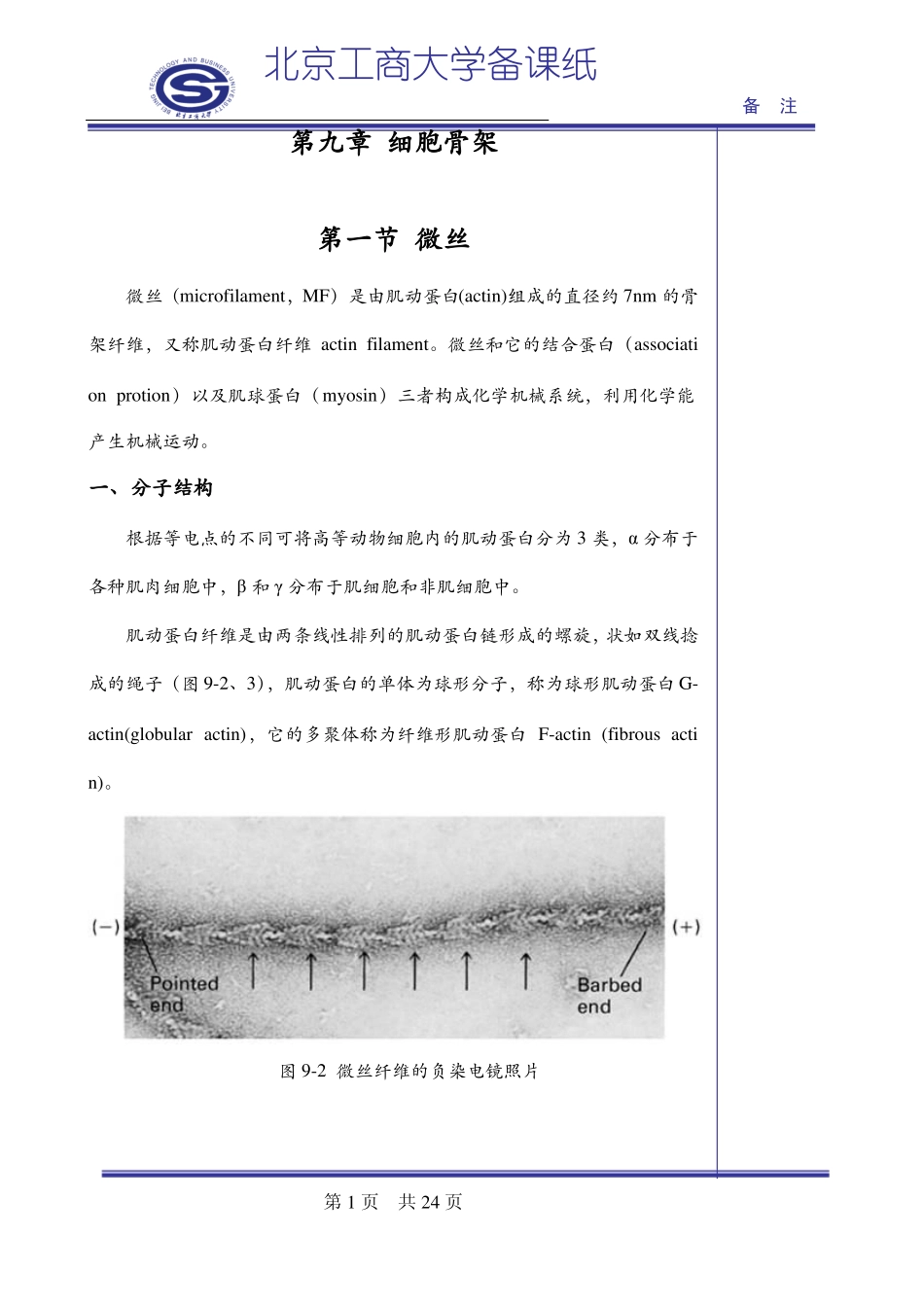
肌动蛋白纤维是由两条线性排列的肌动蛋白链形成的螺旋,状如双线捻成的绳子(图9-2、3),肌动蛋白的单体为球形分子,称为球形肌动蛋白G-actin(globularactin),它的多聚体称为纤维形肌动蛋白F-actin(fibrousactin)
图9-2微丝纤维的负染电镜照片第1页共24页北京工商大学备课纸备注图9-3微丝纤维结构模型肌动蛋白在进化上高度保守,酵母和兔子肌肉的肌动蛋白有88%的同源性
不同类型肌肉细胞的α-肌动蛋白分子一级结构(约400个氨基酸残基)仅相差4~6个氨基酸残基,β-肌动蛋白或γ-肌动蛋白与α-横纹肌肌动蛋白相差约25个氨基酸残基
多数简单的真核生物,如酵母或粘菌,含单个肌动蛋白基因,仅合成一种肌动蛋白
真核生物含有多个肌动蛋白基因,如海胆有11个,网柄菌属(Dictyostelium)有17个,在某些植物中有60个
肌动蛋白要经过翻译后修饰,如N-端乙酰化或组氨酸残基的甲基化
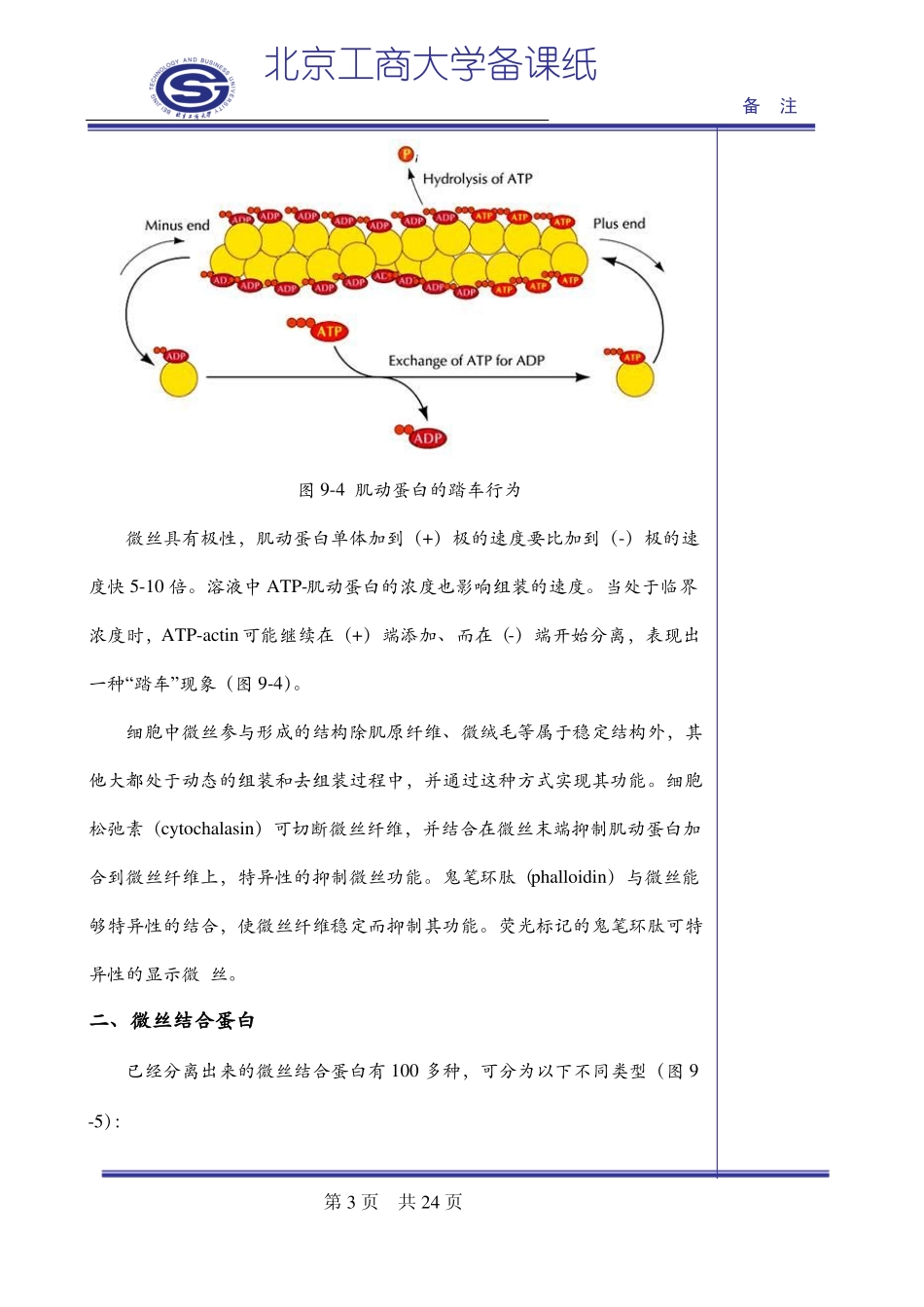
在适宜的温度,存在ATP、K+、Mg2+离子的条件下,肌动蛋白单体可自组装为纤维
ATP-actin(结合ATP的肌动蛋白)对微丝纤维末端的亲和力高,ADP-actin对纤维末端的亲和力低,容易脱落
当溶液中ATP-actin浓度高时,微丝快速生长,在微丝纤维的两端形成ATP-actin“帽子”,这样的微丝有较高的稳定性
伴随着ATP水解