限制性内切酶基础知识‹限制性内切酶限制性内切酶基础知识限制性内切酶关键注意事项限制性内切酶疑难解决指南限制性内切酶在基因组作图与分析中的应用如果没有限制性内切酶,当今的分子生物学研究会是个什么样子
40年来,限制性内切酶这个幕后成员默默推动着许多基础生物学研究和商业化应用
限制性内切酶(又称限制酶)首先是在细菌体内发现的,但后来在部分古细菌中也发现了这种成分
通常,限制性内切酶会切割双链DNA
每个限制性内切酶会识别特定的DNA序列,根据不同的内切酶类型,可在识别序列内或距识别序列不远的位置处切割DNA
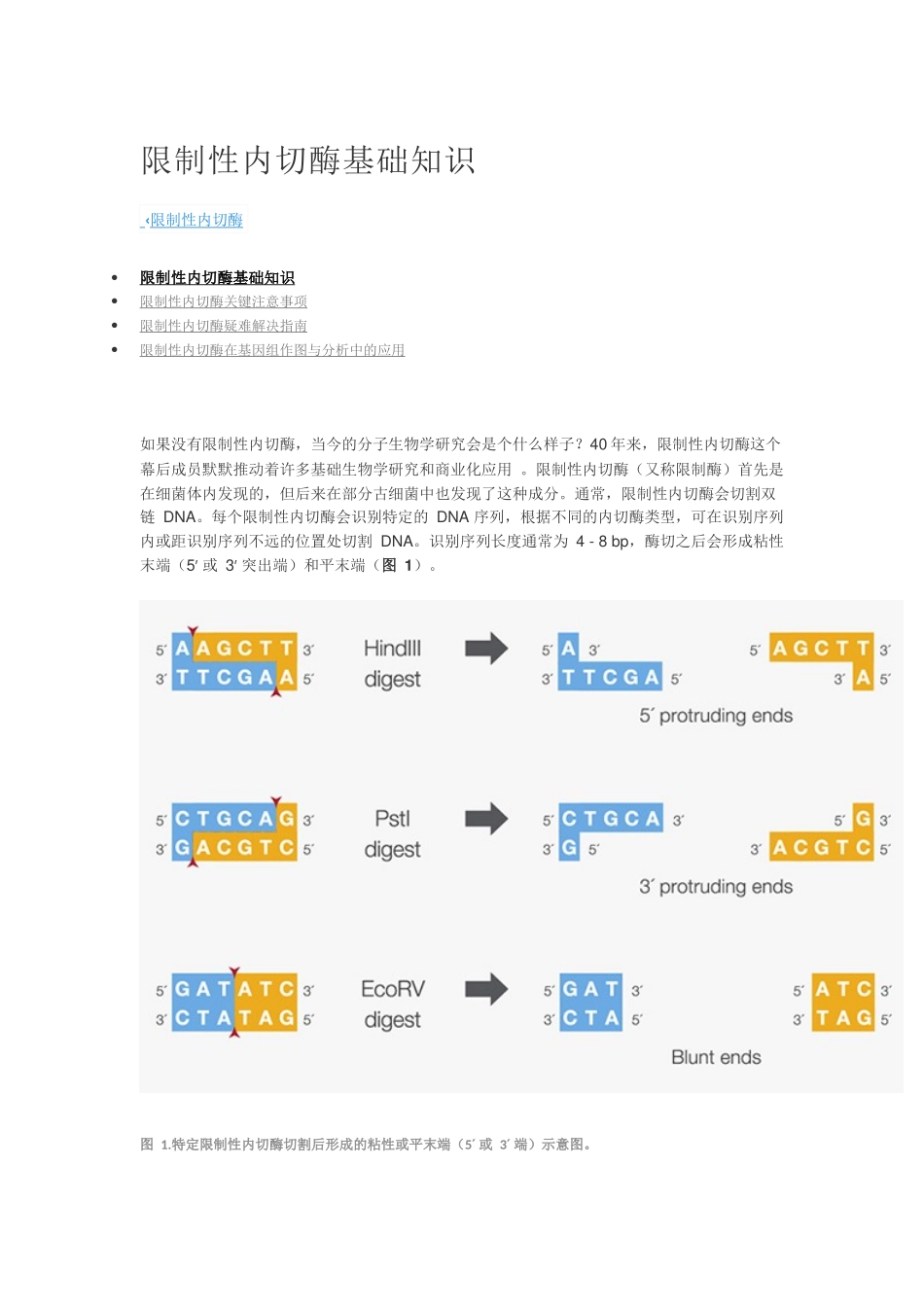
识别序列长度通常为4-8bp,酶切之后会形成粘性末端(5′或3′突出端)和平末端(图1)
特定限制性内切酶切割后形成的粘性或平末端(5′或3′端)示意图
如今业内已经鉴定了大约4000种限制性内切酶,其中超过600种可商业化供应
REBASE是一个非常有用的资源库,可查询限制性内切酶全面信息及更新信息,包括限制性内切酶的特异性、灵敏度和商业化来源等[1]
本页内容:限制性内切酶发展史许多克隆含有空载体限制性内切酶分类识别位点和酶切特异性参考文献限制性内切酶发展史上世纪50年代初期,许多研究团队观测到了噬菌体对于同一物种的不同细菌宿主菌株存在感染效率差异[2,3]
Grasso和Paigen对此的描述如下:使用在一种细菌菌株(例如,大肠杆菌C)内繁殖的噬菌体λ感染同一种类的另一菌株(例如,大肠杆菌K),结果发现,相比于重新感染宿主菌株(大肠杆菌C),大肠杆菌K的感染率出现明显下降
新的宿主(大肠杆菌K)似乎可以选择性抵御或“耐受”侵入的噬菌体
研究人员还发现,这一现象并没有遗传性,因为经过一轮感染后,在新菌株中生长的噬菌体还可以以正常的感染率感染该菌株
这种现象被称为“宿主控制变异”,有关其背后的机制也成为了频繁研究的领域[4]
直到上世纪60年代,人