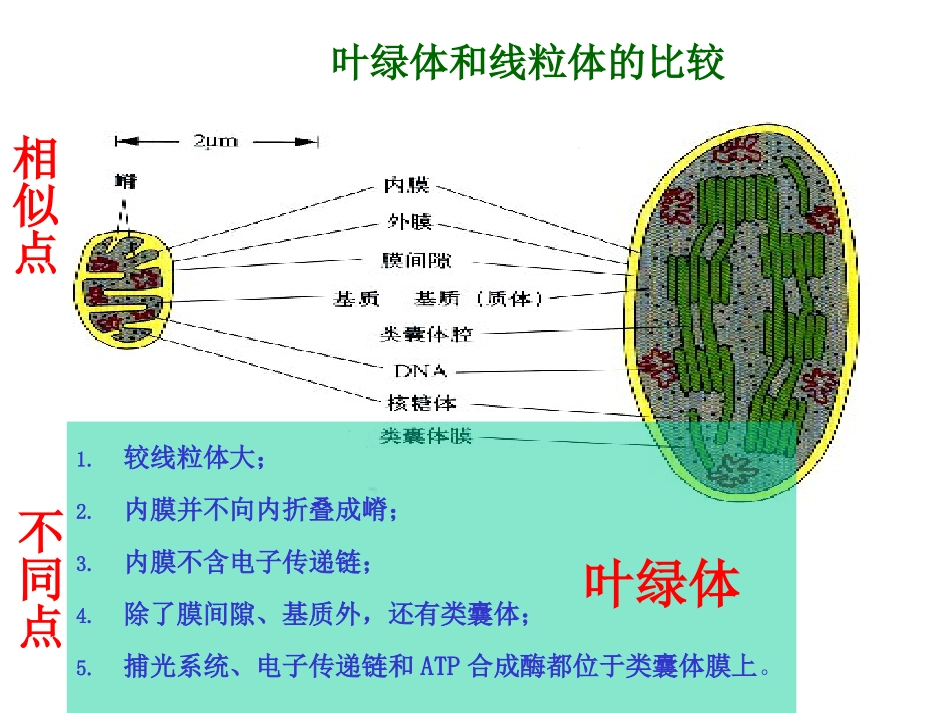
叶绿体和线粒体的比较1
较线粒体大;2
内膜并不向内折叠成嵴;3
内膜不含电子传递链;4
除了膜间隙、基质外,还有类囊体;5
捕光系统、电子传递链和ATP合成酶都位于类囊体膜上
相似点不同点叶绿体第二节线粒体与叶绿体是半自主性的细胞器1
叶绿体与线粒体的DNA2
叶绿体与线粒体的蛋白质合成3
叶绿体与线粒体的蛋白质运送与组装1)线粒体DNA的结构线粒体DNA(简称mtDNA)较小,结构简单,位于线粒体基质中,有时也附着在线粒体内膜上
不含组蛋白,呈环形,与细菌基因组相似
哺乳动物线粒体基因组大小为16500碱基对(bp)
1、叶绿体与线粒体的DNA2)叶绿体DNA1叶绿体DNA(ctDNA)的基本性质和功能性质:高等植物叶绿体DNA(chloroplastDNA,cp或ctDNA)一般是双链环状分子,其周长随种类而不同,大约在40~60µm,含200~2500kbp(千碱基对)
功能:•编码叶绿体rRNA、tRNA和一些核糖体蛋白的基因;•编码PSⅡ、PSI、Cytbf复合体、CF1—CF0ATP合酶和Rubisco等的一些亚单位
1)线粒体基因组编码的RNA和蛋白质线粒体DNA转录的rRNA、tRNA均通过线粒体核糖体用于合成线粒体蛋白质(数目有限)
真核生物线粒体核糖体的RNA组分和蛋白质的构成、大小及对抗生素敏感性方面均不同于细胞质核糖体,但某些方面类似原核生物核糖体
线粒体中的多数蛋白由核基因编码、在细胞质核糖体中合成
故线粒体基因在转录与转译过程中受核基因控制,对核基因具很大依赖性
2、叶绿体与线粒体的蛋白质合成2)参加叶绿体组成的蛋白质来源:•由ctDNA编码,在叶绿体核糖体上合成;•由核DNA编码,在细胞质核糖体上合成;•由核DNA编码,在叶绿体核糖体上合成
1)细胞质中合成的线粒体蛋白质跨膜分选转运至线粒体不同部位:•细胞质核糖体合成的线粒体蛋白