第13章基因组进化的模式基因水平的进化新基因的产生新基因产生的主要方式:1)基因加倍之后的趋异,是新基因产生的主要方式
2)结构域洗牌,即不同的结构域加倍或重组,产生创新功能基因
真核生物约19%的基因产生于外显子洗牌
3)基因裂变与融合,一分为二或者合二为一
原核生物约0
5%的基因由此产生
4)嬗变,由非编码顺序转变为编码顺序(罕见)
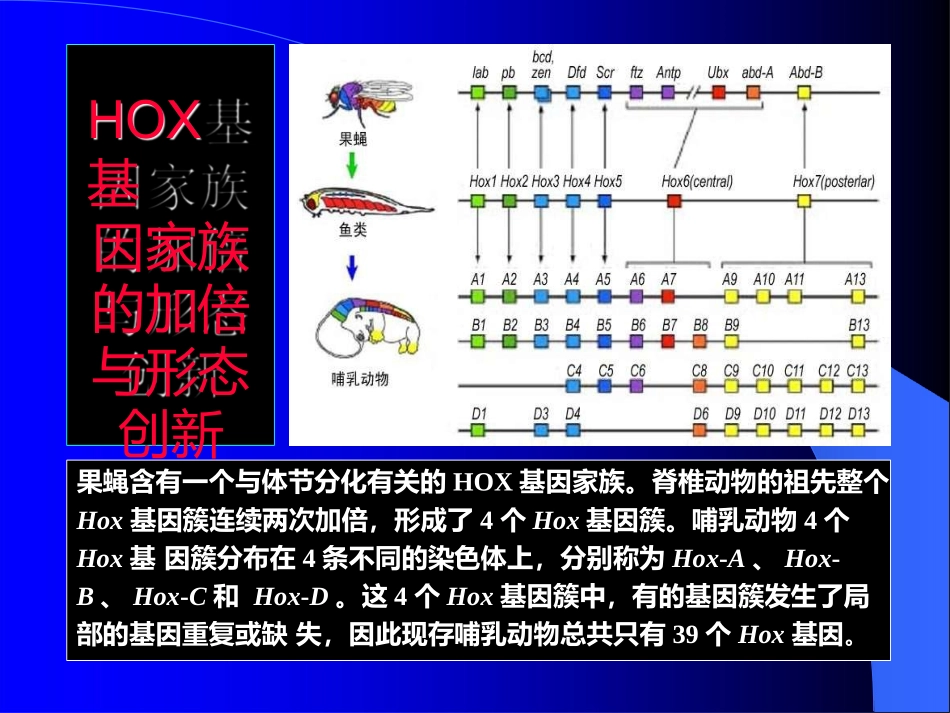
HOX基因家族的加倍与形态创新果蝇含有一个与体节分化有关的HOX基因家族
脊椎动物的祖先整个Hox基因簇连续两次加倍,形成了4个Hox基因簇
哺乳动物4个Hox基因簇分布在4条不同的染色体上,分别称为Hox-A、Hox-B、Hox-C和Hox-D
这4个Hox基因簇中,有的基因簇发生了局部的基因重复或缺失,因此现存哺乳动物总共只有39个Hox基因
Hoxc-6基因在鸡和蛇胚胎中的表达模式Hox基因决定动物个体重复结构的形态,数目及其进化,如像脊椎动物的脊椎骨数目与类别
图左为鸡胚胎发育时期的脊椎组成,Hoxc-6基因控制7数胸椎的模式,蓝色部分将发育为肋骨
图右为乌蛇胚胎,Hoxc-6基因的表达调控使乌蛇胸椎数目(蓝色)极度扩展,从头部一直延伸到尾部
HOX基因簇的进化Hox基因家族在动物的发育与体节模式确定中起着极其重要的作用
哺乳动物的4个Hox基因簇的来源可以追溯到原始的Hox-Evx/Meox基因的加倍,趋异与缺失,代表基因进化的一种主要方式
基因重复与适应性进化-球蛋白基因哺乳动物现存球蛋白基因经历过前后5次加倍与趋异
原和原球蛋白的祖先基因在约5亿年前加倍,在2亿年之后球蛋白和球蛋白祖先基因又出现加倍,随之各自发生分离
现存的肌球蛋白和球蛋白的功能发生了趋异,适应了生物进化的需要:胎儿期表达的球蛋白可满足从母体血液吸收氧的功能,成年表达的球蛋白与氧结合能力减弱
β-球蛋白基因加倍趋异哺乳动物和禽类β-球