干细胞如何衰老和为什么这让我们变老摘要|最近的数据表明,我们的衰老,部分是因为我们干细胞自我更新衰老,如DNA的损伤,以及外在因素,如改变他们的支持niches池
抑制癌症发展的机制,如衰老和凋亡,这依赖于端粒缩短和p53和p16INK4a分子的活动,也可能引起一个有害后果:某些干细胞类型随着年龄复制功能下降
这降低了再生能力似乎导致哺乳动物的某些方面老化,新研究结果指向一个“干细胞假说”引起人类老化症状如虚弱、动脉粥样硬化和2型糖尿病
多样化的收购和遗传因素驱动complexcellular和的哺乳动物的生物老化过程
这个过程似乎是加速环境和行为的因素包括肥胖,糖尿病,终末期肾病和接触诱变剂如化疗,紫外线和吸烟
同样的,各种遗传病变改变DNA代谢和核结构伴随着人类早衰综合征如共济失调毛细血管扩张症,沃纳综合症和哈钦森-德卡尔曼氏综合症
最引人注目的在酵母、果蝇,黑腹菌属,秀丽隐杆线虫和老鼠的基因研究建立了至关重要作用的是insulin-insulinlike生长因子-1(IGF1)-AKT-forkhead转录因素类经典0FOXO)途径和氧自由基监管机构作为决定因素的寿命在这些模型系统
尽管这些不同的实验观察显示相同分子主题,人类衰老的生理条件仍然一个谜
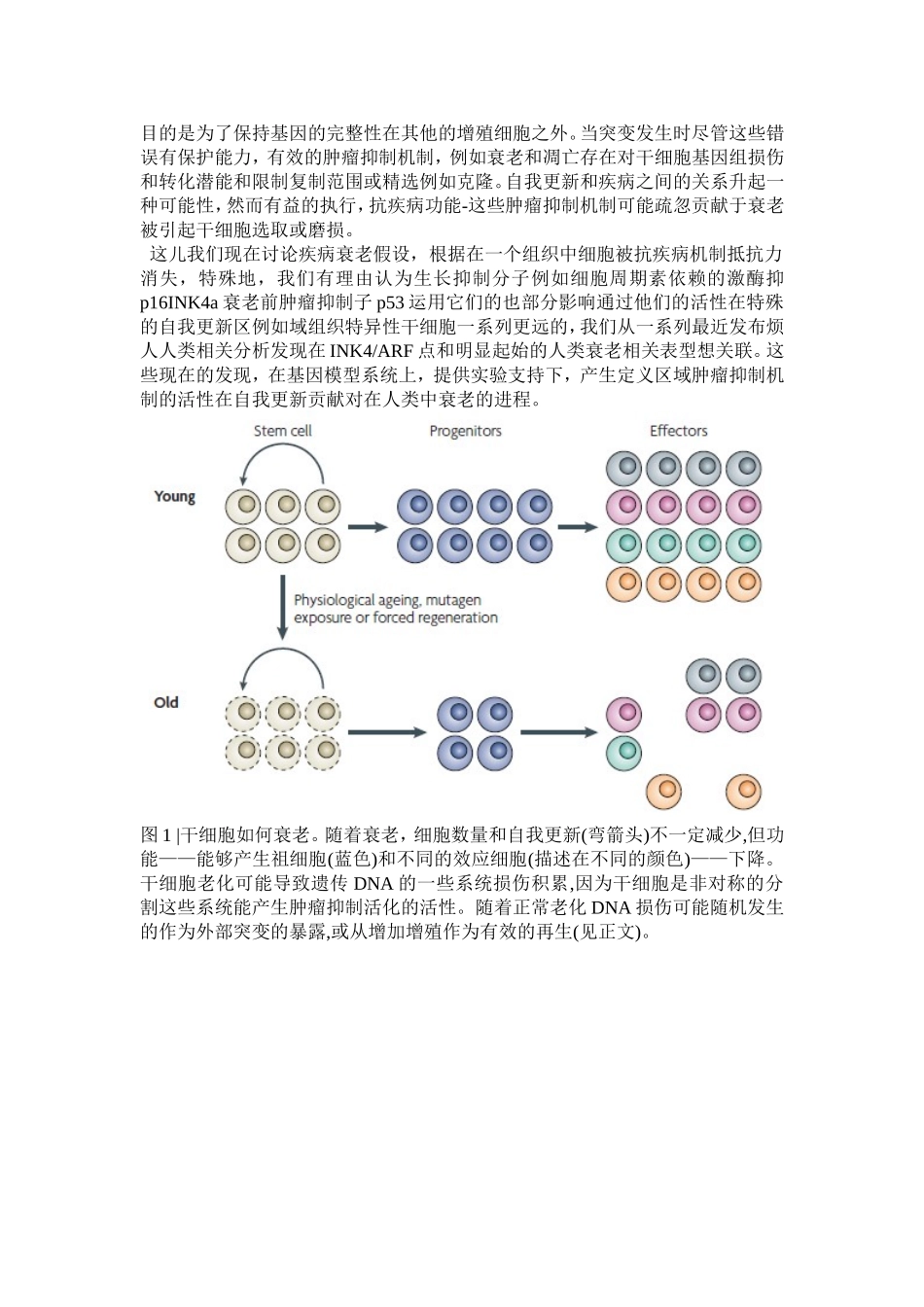
在此综述中,我们认为哺乳动物衰老结果的某些方面从一个老化引起的复制功能减少的某些组织特异性再生细胞-成人干细胞(图1)
这些罕见的和专一细胞都需要组织替换贯穿在整个人类的寿命,和似乎具有一些特定的生理和生化特性,特别是自我更新的能力
最近的证据支持这个模型,干细胞在一些组织在很大程度上处于一个静止的状态但是,即使经过长时间沉默仍可以巧妙回到细胞周期响应到细胞外的信号
一旦刺激分化,产生干细胞数不分化后代,进而通过随后的增殖循环产生不同的效应细胞(框1)
这种“分层”分化计划的意义在于揭示生物的长寿前景-它允许生产大分化细